Классификация ВПР
Согласно международной классификации (ВОЗ) врожденные пороки развития подразделяют:
1) по частоте: на распространенные – 1: 1000 и более, умеренно частые –
0,1-0,99 на -1000 и очень редкие – меньше 0,01 на 1000;
2) по степени тяжести и прогнозу для жизнеспособности: на летальные – смерть до репродуктивного периода (0,6%), средней степени тяжести – не угрожают жизни, но требуют оперативного вмешательства (1,9-2,5%) и
малые аномалии развития или информативные морфологические варианты
(3,5%); 3) по проявлению: на изолированные (монотопический дефект поля,
политопический дефект поля и sequence), множественные (синдром,
ассоциация, случайныекомбинации);
4) по этиологии: на моногенные – 6%, хромосомные –5%, внешнесредовые
(тератогены, материнские факторы) – 6%, мультифакториальные – 63%,
неустановленные причины –20%.
В зависимости от последовательности возникновения (типа «ошибок») в
процессе морфогенеза пороки развития делят на 4 группы (Опитц Дж., 1977,
Лазюк Г.И., 1983, Козлова С.И., 1986):
—мальформация (malformation) – морфологический дефект органа, части органа или большого участка тела в результате внутреннего нарушения процесса развития (генетически детерминированный процесс);
—дизрупция (disruption)- морфологический дефект органа, части органа или большого участка тела в результате внешнего препятствия или какого-либо воздействия на изначально нормальный процесс развития (тератогенные факторы и нарушение имплантации);
—деформация (deformation) –нарушение формы, вида или положения части тела, обусловленное механическими воздействиями;
—дисплазия (dysplasia)- нарушенная организация клеток в ткани (тканях) и ее морфологический результат (процесс и следствие дисгистогенеза).
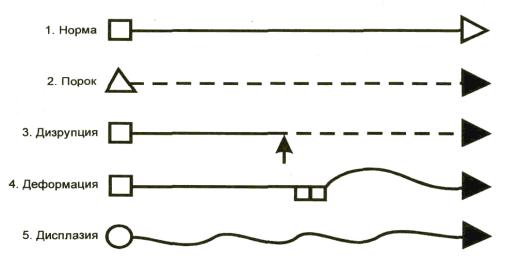
Рис. 9. Схема разных типов ошибок морфогенеза (J. Opitz, 1977).
В зависимости от времени и объекта воздействия вредных факторов ВПР делятся на четыре типа: гаметопатии – связаны с возникновением мутаций в половых клетках родителей (генные, хромосомные синдромы);
бластопатии – поражение бластоцисты (зародыш первых 15 дней после оплодотворения, следствие бластопатий – двойниковые пороки, циклопия,
сиреномелия, пустые зародышевые мешки); эмбриопатии – возникают в период от 16 дня до конца 8 недели беременности (в этот период происходит активный органогенез и гистогенез, большинство пороков образуется в этот период); фетопатии – повреждение плода в период от 9 недели до окончания беременности (крипторхизм, тазовая почка, метаболические фетопатии). Ниже мы приводим описание наиболее известных эмбриофетопатий.
Фетальный вальпроевый синдром
Данный синдром обусловлен приемом во время беременности противосудорожных препаратов вальпроевой кислоты. Дети рождаются с дефицитом массы и длины тела. Типичны аномалии лица: небольшие орбиты, гипоплазия средней части лица, короткий вздернутый нос, длинный уплощенный фильтр, широкая переносица, эпикант, микрогнатия, маленький рот, расщелина губы, расщелина неба, микротия, анотия. Также для
синдрома характерны аномалии черепа: тригоноцефалия, узкий бифронтальный диаметр, синостоз швов. К другим проявлениям синдрома относятся полидактилия, дефекты нервной трубки (спинномозговая грыжа),
пороки сердца (ДМЖП, ОАП, коарктация аорты), гипоспадия, ВПР ЖКТ. У
детей наблюдается отставание в статико-моторном и умственном развитии.
Фетальный синдром краснухи (синдром Грега)
Впервые синдром описан в 1941 году N. Gregg. Риск для плода у серопозитивных женщин составляет 40-90%. Инфицирование плода вирусом краснухи может вызвать спонтанный аборт, преждевременные роды.
Типичны поражения сердца, органов зрения и слуха. Среди ВПР глаз отмечаются катаракта, микрофтальмия, пигментная ретинопатия, глаукома,
миопия, страбизм, колобома. Постоянный признак – нейросенсорная глухота
(одноили двусторонняя). Из ВПР сердца встречаются открытый аортальный проток, стеноз легочной артерии, дефект межжелудочковой перегородки. Характерно выделение вируса из различных тканей, выявление специфических антител в сыворотке плода. У больных детей наблюдается умственная отсталость, дефицит роста, микроцефалия, гидроцефалия,
пренатальная гипоплазия, иммунологический дефицит, персистенция вируса во многих органах, аномалии зубов, гемолитическая анемия. Основным видом профилактики является иммунизация девочек по общепринятой схеме.
Фетальный цитомегаловирусный синдром
Высокий риск развития данного синдрома связан с первичной цитомегаловирусной инфекцией в первые 6 месяцев гестации. У 10% плодов имеются тяжелые ВПР. Характерны внутриутробная задержка развития,
гепатоспленомегалия, желтуха, инфицирование плода цитомегаловирусом.
Дети рождаются недоношенными, с низкой массой тела. Нередко отмечаются микроцефалия, перивентрикулярные кальцификаты (энцефалит),
гидроцефалия, умственная отсталость, гипотония, судороги. Возможно поражение глаз: страбизм, хореоретинит, микрофтальмия, катаракта,
кальцификаты в сетчатке. Часто у детей выявляется тромбоцитопения,
гемолитическая анемия, петехии. Профилактика: избегать контактов,
пренатальная диагностика (УЗИ) – вентрикуломегалия, кальцификаты в головном мозге.
Фетальный алкогольный синдром
(FAS, алкогольная эмбриофетопатия)
Частота ФАС в США — от 0,2 до 2 на 1000. При хроническом алкоголизме матери риск развития ФАС составляет 25-45%. У детей отмечается дефицит роста и веса. Характерно поражение центральной нервной системы: тремор,
судороги, опистотонус, гипотония, нарушения поведения, умственная отсталость, гидроцефалия, микроцефалия. Типичны изменения лица:
короткая глазная щель, сглаженный губной желобок, тонкая верхняя губа,
блефарофимоз, эпикант, птоз, косоглазие, удлиненное лицо, микрогения. У
детей выявляются аномалии пальцев, дисплазия ТБС, деформации грудной клетки, гипоспадия, крипторхизм, удвоение влагалища, ВПР сердца (чаще ДМЖП), ангиомы, фиброз печени.
В медицинской практике также широко используется классификация аномалий развития, которая основана на органно-системном (анатомо-
физиологическом) принципе.
К распространенным видам пороков в практике врача любой специальности относятся следующие:
—агенезия – полное врожденное отсутствие органа;
—аплазия – врожденное отсутствие органа с сохранением его сосудистой ножки;
—атрезия и стеноз – соответственно полное отсутствие или сужение канала
(отверстия);
— врожденная гипоплазия – недоразвитие органа, проявляющееся дефицитом относительной массы (отношение абсолютной массы органа к абсолютной массе тела плода или ребенка, выраженное в процентах) или размеров органа, превышающее отклонение от средних показателей в две сигмы
(средняя ошибка) для данного возраста: простая – без нарушения структуры и диспластическая – с нарушением структуры;
— врожденная гипотрофия – уменьшение массы тела плода или новорож-
денного;
— врожденная гипертрофии (гиперплазия) – увеличение относительной мас-
сы (размеров) органа за счет увеличения объема (гипертрофия) или количества (гиперплазия) клеток;
— гетероплазия – нарушение дифференцировки отдельных типов тканей,
например, клетки плоского эпителия пищевода в дивертикуле Меккеля;
—гетеротопия – наличие клеток, тканей или участка органа в другом органе или в тех зонах органа, где в норме их не должно быть;
—эктопия – смещение органа в нетипичное для него место;
—дизрафия или арафия – незаращение эмбриональной щели (расщелина губы или неба, позвоночника);
—дисхрония – нарушение темпов развития (ускоренное или замедленное);
—макросомия (гигантизм) – увеличение длины тела;
—неразделение (слияние) – может быть органа или монозиготных близнецов
(краниопаги, торокопаги, сросшиеся черепами или грудными клетками близнецы);
— персистирование – сохранение эмбриональных структур, в норме исчеза-
ющих к определенному периоду онтогенеза, например, незаращение баталлова протока или овального окна у ребенка в возрасте старше трех месяце;
— нарушение лобуляции – дополнительная доля легкого, печени;
—инверсия-зеркальноерасположениеорганов:
—удвоение органов;
—образование водянок – гидроцефалия, гидронефроз.
Для объяснения этиологических и патогенетических связей между разными типами аномалий развития предложены следующие понятия:
следствие, синдром, ассоциация.
Синдром – комплекс множественных пороков и аномалий развития,
патогенетически и этиологически связанных между собой; это устойчивое сочетание двух и более не индуцированных друг другом пороков развития в разных системах организма.
Следствие (аномалад, секвенция, вторичный порок) – комплекс пороков развития, возникающий вследствие действия одного ВПР или физического фактора, который вызывает каскад вторичных нарушений (связь между пороками патогенетическая). Например, спорадический аномалад Пьера-
Робена, при котором первичным пороком является микрогения, а его следствием будут уменьшение ротовой полости, глоссоптоз, расщелина неба.
Ассоциация – неслучайное сочетание нескольких пороков и аномалий развития, наблюдающееся у 2-х и более пациентов, не известных как синдром или следствие. Например, ассоциация OEIS – омфалоцеле,
экстрофия мочевого пузыря, атрезия заднего прохода, дефекты позвоночника; ассоциация VAСTERL (MIM:192350) – атрезия пищевода и заднего прохода, трахео-эзофагальные фистулы, пороки почек, сердца,
дефекты позвоночника, радиальные дисплазии.
В последнее время внимание врачей различных специальностей обращается на диагностическое значение малых аномалий развития. Малая аномалия развития (МАР, дисгенезия, информативный морфологический вариант, дизэмбриологический признак, дисморфологическая черта,
микропризнак, микродегенеративный признак и др.) – это стойкое морфологическое нарушение органа, участка тела, не выходящее за пределы вариаций или находящееся у крайних границ вариаций строения и не сопровождающееся нарушением его функции. Известно около 200 МАР.
Малые аномалии развития с разной частотой распространены в популяции в рамках нормальной вариабельности. Например, сросшиеся брови,
поперечная складка ладони, гипертелоризм, деформация ушных раковин,
клинодактилия V пальца, гипертрихоз и др. Микропризнаки – это морфологические нарушения на конечной тонко настроенной стадии
развития, т.е. нарушения гистогенеза. Для них характерна стабильность – имеются при рождении, не исчезают с возрастом (исключение капиллярная гемангиома, эпикант, пигментные пятна и др.). Малые аномалии не имеют серьезного медицинского или косметического значения, но нередко выступают как значащие симптомы наследственной патологии. Выявление данных изменений фенотипа ребенка требует динамического наблюдения за ним и проведения цитогенетического и ультразвукового исследований.
В диагностических целях следует учитывать 3 важных параметра МАР: их количество, сочетание, «качество». При наличии 3-5 и более микропризнаков
часто выявляются большие аномалии развития или нарушения интеллекта. В
качестве маркеров некоторых наследственных заболеваний и синдромов МВПР могут рассматриваться комплексы из 4, 5-и и более МАР. Например,
полидактилия, гипертелоризм глаз и сосков, дополнительные соски – эти
«малые» аномалии предполагают наличие у пациента пороков мочевыводящей системы. Большое значение имеет сочетание МАР.
Например, для синдрома Дауна характерно наличие монголоидного разреза глаз, эпиканта, брахидактилии, клинодактилии, сандалевидной щели,
поперечной ладонной складки. Наибольшее диагностическое значение имеет
«качество» МАР, т.е. выделение таких признаков, которые среди здоровых пациентов почти не встречаются: «крыловидные» складки шеи
(с.Шерешевского-Тернера, с.Нунан), постаксиальная полидактилия (с.Барде-
Бидля), вертикальные насечки на мочке уха (с.Беквита-Видемана).
Наибольшей значимостью обладают такие микропризнаки, которые встречаются редко и среди больных с наследственной и врожденной патологией, и в популяции здоровых людей. Например, диастема (частота в популяции 15%) и гиподонтия (очень редко встречающийся признак) несут разную диагностическую нагрузку. Отдельные микропризнаки встречаются чаще у недоношенных и маловесных детей. Они могут быть этническими особенностями, например, эпикант является нормой у монголоидной расы –
60-65%. МАР можно разделить на 3 группы: альтернативные, измерительные
Схема разных типов ошибок морфогенеза — Справочник по медицине PRO7
Review
. 1998 Apr 1;76(4):291-6.
Affiliations
-
PMID:
9545092
Review
Errors of morphogenesis and developmental field theory
M L Martínez-Frías et al.
Am J Med Genet.
.
Abstract
Field theory provides a rational basis for birth defects terminology. During blastogenesis in higher metazoa, pattern formation in the primary field leads to the establishment of upstream expression domains of growth and transcription factors, which, in various permutations and at specific sites and times, lay down the pattern of progenitor fields. Further spatially coordinated, temporally synchronized, and epimorphically hierarchical morphogenetic events, mostly during organogenesis, lead to the attainment of final form in the secondary, epimorphic fields. Because of shared molecular determinants, spatial contiguity, and close timing of morphogenetic events during blastogenesis, most malformations arising during blastogenesis are polytopic, i.e., involving two or more progenitor fields, e.g., acrorenal, cardiomelic, gastromelic, or splenomelic anomalies. Defects of organogenesis tend to be monotopic malformations, e.g., cleft palate or postaxial polydactyly. We suggest that what were called «associations» (e.g., VATER, schisis) be designated primary polytopic developmental field defects, or simply polytopic field defects, and that the term «association» be reserved for the original definition of a statistical combination of anomalies (mostly of organogenesis) [Spranger et al. (1982): J Pediatr 100:160-165]. If genetically caused or predisposed, all structures involved in a polytopic or monotopic malformation are genetically abnormal, whereas the parts secondarily affected as a consequence of a malformation sequence (e.g., spina bifida) are genetically normal. Polytopic field anomalies, per se, must be distinguished from pleiotropy, although such anomalies may constitute a part of pleiotropy (e.g., in trisomy 18). Because they are downstream from pattern-forming events in the primary field, multiple anomalies of organogenesis more likely represent syndromal pleiotropy.
Similar articles
-
[Developmental abnormalities in humans].
Opitz JM.
Opitz JM.
Monatsschr Kinderheilkd. 1991 May;139(5):259-72.
Monatsschr Kinderheilkd. 1991.PMID: 1870596
Review.
German. -
Limb anomalies from evolutionary, developmental, and genetic perspectives.
Opitz JM.
Opitz JM.
Birth Defects Orig Artic Ser. 1996;30(1):35-77.
Birth Defects Orig Artic Ser. 1996.PMID: 9125339
Review.
-
Pathogenetic classification of a series of 27,145 consecutive infants with congenital defects.
Martínez-Frías ML, Bermejo E, Frías JL.
Martínez-Frías ML, et al.
Am J Med Genet. 2000 Jan 31;90(3):246-9.
Am J Med Genet. 2000.PMID: 10678664
Review.
-
Primary developmental field. III: Clinical and epidemiological study of blastogenetic anomalies and their relationship to different MCA patterns.
Martínez-Frías ML, Frías JL.
Martínez-Frías ML, et al.
Am J Med Genet. 1997 May 2;70(1):11-5. doi: 10.1002/(sici)1096-8628(19970502)70:1<11::aid-ajmg3>3.0.co;2-u.
Am J Med Genet. 1997.PMID: 9129734
-
The developmental field concept.
Opitz JM.
Opitz JM.
Am J Med Genet. 1985 May;21(1):1-11. doi: 10.1002/ajmg.1320210102.
Am J Med Genet. 1985.PMID: 4003434
Cited by
-
Patterns of co-occurring birth defects in children with anotia and microtia.
Schraw JM, Benjamin RH, Shumate CJ, Canfield MA, Scott DA, McLean SD, Northrup H, Scheuerle AE, Schaaf CP, Ray JW, Chen H, Agopian AJ, Lupo PJ.
Schraw JM, et al.
Am J Med Genet A. 2023 Mar;191(3):805-812. doi: 10.1002/ajmg.a.63081. Epub 2022 Dec 21.
Am J Med Genet A. 2023.PMID: 36541232
-
Anorectal Malformations in Male Monozygotic Twins and Review of Literature.
Kundal VK, Kundal R, Meena AK, Debnath PR, Shukla AK.
Kundal VK, et al.
J Indian Assoc Pediatr Surg. 2022 Mar-Apr;27(2):136-139. doi: 10.4103/jiaps.JIAPS_344_20. Epub 2022 Mar 1.
J Indian Assoc Pediatr Surg. 2022.PMID: 35937120
Free PMC article. -
Five-year experience of anorectal malformation with oesophageal atresia in tertiary care hospital.
Chowdhary S, Panigrahi P, Kumar R.
Chowdhary S, et al.
Afr J Paediatr Surg. 2020 Jul-Dec;17(3 & 4):49-53. doi: 10.4103/ajps.AJPS_54_17.
Afr J Paediatr Surg. 2020.PMID: 33342833
Free PMC article. -
Maternal drug use and the risk of anorectal malformations: systematic review and meta-analysis.
Zwink N, Jenetzky E.
Zwink N, et al.
Orphanet J Rare Dis. 2018 May 10;13(1):75. doi: 10.1186/s13023-018-0789-3.
Orphanet J Rare Dis. 2018.PMID: 29747656
Free PMC article. -
Full Spectrum of VACTERL in New Born.
Harjai MM, Holla RG, Kale R.
Harjai MM, et al.
Med J Armed Forces India. 2008 Jan;64(1):84-5. doi: 10.1016/S0377-1237(08)80163-3. Epub 2011 Jul 21.
Med J Armed Forces India. 2008.PMID: 27408093
Free PMC article.No abstract available.
Publication types
MeSH terms
LinkOut — more resources
-
Full Text Sources
- Wiley
-
Medical
- MedlinePlus Health Information
Глава 1. Нерешённые проблемы биологии
1.1. Предпосылки успеха
Цель механистического подхода в биологическом исследовании особенно ясно была выражена более ста лет назад Томасом Хаксли в следующем определении: «Зоологическая физиология — это доктрина функций или действий животных. Она рассматривает тела животных как машины, побуждаемые к действию различными силами и совершающие некоторое количество работы, которую можно выразить на языке обычных сил природы. Конечная цель физиологии состоит в том, чтобы вывести факты морфологии, с одной стороны, и факты экологические — с другой, из законов молекулярных сил вещества»2.
Эти идеи прослеживаются во всём последующем развитии физиологии, биохимии, биофизики, генетики и молекулярной биологии. Эти науки во многих отношениях достигли блестящих успехов и более всех молекулярная биология. Открытие структуры ДНК, «разгадка генетического кода» и выяснение механизма синтеза белка являются впечатляющими подтверждениями применимости механистического подхода.
Наиболее ярко выраженными и влиятельными из современных сторонников механистической теории являются представители молекулярной биологии. Своё изложение теории они обычно начинают с категорического отрицания виталистических3 и организмических теорий. Их называют пережитками «примитивных» верований, которые обречены отступать всё дальше и дальше, по мере продвижения механистической биологии. Затем рассуждения строятся следующим образом4:
Химическая природа генетического материала, ДНК, теперь известна, как и генетический код, определяющий последовательность аминокислот в белках. Механизм синтеза белка также понят достаточно глубоко. Сейчас раскрыта структура многих белков. Все ферменты — белки, и они катализируют сложные цепи и циклы биохимических реакций, которые образуют метаболизм организма. Метаболизм контролируется биохимической обратной связью; известно несколько механизмов, которые могут регулировать скорость ферментативных реакций. Белки и нуклеиновые кислоты самопроизвольно агрегируют с образованием таких структур, как вирусы и рибосомы. Если набор свойств белков, а также свойств других физико-химических систем, таких как липидные мембраны, будет известен, свойства живых клеток могут быть, в принципе, полностью объяснены.
Ключом к решению проблем дифференциации и развития, о которых пока известно очень мало, является понимание механизма контроля синтеза белка. Способ контроля синтеза некоторых метаболических ферментов и других белков детально установлен для бактерии Escherichia coli. В высших организмах контроль синтеза белка осуществляется с помощью более сложных механизмов, но они скоро должны быть раскрыты. Так что дифференциация и развитие должны быть объяснимы с помощью серий химически управляемых «переключателей», которые «включают» и «выключают» отдельные гены или их группы.
Способ приспособления частей живых организмов к функциям целого, а также явная целенаправленность структуры и поведения организмов, могут быть объяснены случайными генетическими мутациями, за которыми следует естественный отбор, причём отбираются гены, увеличивающие способность организма выживать и воспроизводить; вредные мутации исключаются. Так неодарвинистская теория эволюции пытается объяснить целенаправленность; совершенно необязательно предполагать, что она связана с какими-либо таинственными «витальными факторами».
Очень мало известно о функционировании центральной нервной системы, но со временем, благодаря успехам биохимии, биофизики и электрофизиологии, станет возможным объяснить то, что мы называем разумом, на механистическом языке физико-химических процессов в мозге. Таким образом, живые организмы в принципе полностью объяснимы в терминах физики и химии. То, что мы сегодня не понимаем в механизмах развития и в деятельности центральной нервной системы, является следствием невероятной сложности этих проблем; но теперь, вооруженные новыми мощными концепциями молекулярной биологии и компьютерными моделями, мы можем взяться за решение этих задач в таких масштабах и такими способами, которые раньше были недоступны.
* * *
В свете прошлых успехов вполне понятен оптимизм и уверенность в том, что все проблемы биологии могут быть до конца решены с позиций механистического подхода. Но реалистическая оценка перспектив механистического объяснения должна опираться на нечто более серьезное, нежели историческая экстраполяция; такая оценка может быть сформирована только после рассмотрения важнейших проблем биологии и способов, которыми они предположительно могут быть решены.
1.2. Проблемы морфогенеза
Биологический морфогенез можно определить как «появление характерной и специфической формы в живых организмах»5. Первая проблема заключается именно в том, что форма обретает существование. Биологическое развитие является эпигенетическим, если появляются новые структуры, которые не могут быть объяснены как результат развёртывания или роста структур, уже присутствующих в яйцеклетке в начале развития.
Вторая проблема состоит в том, что многие развивающиеся системы обладают способностью к регуляции; другими словами, если часть развивающейся системы удаляется (или если добавляется дополнительная часть), система продолжает развиваться, и образуется более или менее нормальная структура. Классическая демонстрация этого явления была проведена и 1890-е годы Г.Дришем в его экспериментах на эмбрионах морского ежа. Когда одну из клеток очень молодого эмбриона на двухклеточной стадии развития убивали, из оставшейся клетки развивалась не половина морского ежа, но совершенно целый морской ёж, хотя и меньшего размера. Точно также, маленькие, но целые организмы развивались после разрушения любых одной, двух или трёх клеток эмбриона на четырёхклеточной стадии. Напротив, после слияния двух молодых эмбрионов развивался один гигантский морской ёж6.
Регуляция была продемонстрирована на многих развивающихся системах. Однако в процессе развития организма эта способность утрачивается, когда определяется судьба его отдельных частей. Но даже в системах, где эта детерминированность имеет место на ранней стадии, например, в эмбрионах насекомых, регуляция может осуществляться после повреждения яйца (рис.1).
Рис.1. Пример регуляции. Слева — нормальный эмбрион стрекозы Platycnemis pennipes. Справа — небольшой, но полноценный эмбрион, полученный из задней части яйца, перетянутого в средине вскоре после его откладывания (По Weiss, 1942)
_20.03.2002/novaya_nauka_o_zhizni_chast_2/image001.jpg "Новая наука о жизни (часть 2)")
Подобные результаты показывают, что развивающиеся системы движутся к морфологической цели и обладают некоторым свойством, которое определяет эту цель и позволяет им достичь её, даже если части системы удалены и созданы препятствия для нормального развития.
Третья проблема — регенерация, то есть способность организмов заменять или восстанавливать повреждённые структуры. Растения обнаруживают удивительно широкий диапазон возможностей регенерации, и то же можно сказать о низших животных; например, если плоского червя разрезать на несколько кусочков, из каждого может вырасти целый червь. Даже многие позвоночные обладают поразительными способностями к регенерации; например, если хирургическим путём удалить линзу из глаза тритона, из края радужной оболочки вырастает новая линза (рис.2); при нормальном эмбриональном развитии линза образуется совершенно другим путём, из клеток кожи. Этот тип регенерации был впервые обнаружен Г.Вольфом, который умышленно выбрал такой вид повреждения организма, который не мог случайно произойти в природе; поэтому такой регенеративный процесс не мог быть связан с естественным отбором7
Рис.2. Регенерация линз из границ радужной оболочки глаза тритона после хирургического удаления природной линзы (Cf. Needham, 1942)
_20.03.2002/novaya_nauka_o_zhizni_chast_2/image002.jpg "Новая наука о жизни (часть 2)")
Четвёртая проблема — просто сам факт воспроизведения: отдельная часть родительского организма становится новым организмом; часть становится целым.
* * *
Все эти явления могут быть понятны, только если допустить наличие причинных сущностей, которые составляют нечто большее, чем сумма частей развивающихся систем, и определяют цели процессов развития.
Виталисты приписывают эти свойства витальным факторам, органицисты — морфогенетическим полям, механицисты — генетическим программам.
Концепция генетических программ основана на аналогии с программами, которые определяют работу компьютеров. Она предполагает, что оплодотворённое яйцо содержит предварительно сформированную программу, которая каким-то образом определяет морфогенетические задачи организма, а также координирует и контролирует его развитие в направлении, соответствующем этим задачам. Но генетическая программа должна включать нечто большее, чем химическую структуру ДНК, поскольку идентичные копии ДНК передаются всем клеткам; если бы все клетки были запрограммированы одинаково, они не могли бы развиваться различным образом. Тогда, что же такое это нечто? Чтобы ответить на этот вопрос, основополагающую идею приходится представлять в виде отдельных неопределённых предположений о физико-химических взаимодействиях, каким-то образом структурированных во времени и пространстве; иначе говоря, проблема лишь предстаёт в иной формулировке8.
Существует и другое серьёзное затруднение. Компьютерная программа закладывается в компьютер разумным сознательным существом — программистом. Она создаётся и записывается для достижения определённой цели.
Если генетическую программу рассматривать как аналог компьютерной программы, то тогда должна существовать некая целенаправленно действующая сущность, исполняющая роль программиста. Но если предположить, что генетические программы аналогичны не обычным компьютерным программам, а программам самовоспроизводящихся, самоорганизующихся компьютеров, то их-то пока просто не существует. А если бы даже они существовали, то должны были бы быть невероятно сложным образом запрограммированы теми, кто их изобрёл. Единственный выход из этой дилеммы — утверждать, что генетическая программа построена в ходе эволюции путём сочетания случайных мутаций и естественного отбора. Но тогда подобие с компьютерной программой исчезает, и эта аналогия становится бессмысленной.
Ортодоксальные механицисты отвергают идею, что кажущееся целенаправленным поведение развивающихся, регулирующих и регенерирующих организмов указывает на присутствие витального фактора, который ведёт их к достижению морфологических целей. Но до тех пор, пока механистические объяснения зависят от телеологических концепций, таких как генетические программы или генетические инструкции, целенаправленность может быть объяснена только тем, что она была заложена в них ранее. Действительно, свойства, приписываемые генетическим программам, удивительно похожи на те, которыми виталисты наделяют свои гипотетические витальные факторы; ирония в том, что генетические программы оказываются весьма похожими на витальный фактор в механистическом облике9.
Конечно, тот факт, что биологический морфогенез в настоящее время не может быть объяснён строго механистически, не доказывает, что он не может быть так объяснён никогда. Перспективы подобного объяснения рассматриваются в следующей главе. Но на сегодняшний день убедительный ответ с механистических позиций дать невозможно.
1.3. Поведение
Если проблемы морфогенеза пугающе сложны, то ещё более трудноразрешимы проблемы поведения. Во-первых, инстинкт. Обратите внимание, например, как пауки могут плести паутину, не учась этому у других пауков’10 . Или посмотрите, как ведут себя европейские кукушки. Птенцов выводят и воспитывают птицы других видов, и молодые кукушата никогда не видят своих родителей. В конце лета взрослые кукушки летят к своим зимним гнездовьям в Южной Африке. Примерно через месяц молодые кукушки собираются в стаи и также летят в соответствующий район Южной Африки, где они присоединяются к своим старшим собратьям11. Они инстинктивно знают, что должны мигрировать и когда именно, узнают друг друга, собираясь в стаи, и понимают, в каком направлении надо лететь и где их место назначения.
Во-вторых, существует проблема, связанная с многочисленными примерами регуляции поведения, в которых более или менее нормальный результат достигается несмотря на изменения в тех или иных частях биологической системы. Например, собака после ампутации ноги регулирует свою двигательную активность так, что она может ходить на трёх лапах. Другая собака после удаления части одного из полушарий мозга постепенно восстанавливает большинство из своих прежних способностей. Третья — встречает препятствия, произвольно поставленные на её пути. Но все три собаки могут пройти от одного места до другого, к которому они хотят добраться, несмотря на повреждения их двигательных органов, центральной нервной системы или изменения окружения.
В-третьих, проблема, возникающая в связи с обучением и разумным поведением; появляется новый способ поведения, который, очевидно, не может быть полностью объяснён ранее перечисленными причинами.
Гигантская пропасть незнания лежит между всеми этими феноменами и установленными фактами молекулярной биологии, биохимии, генетики, нейрофизиологии. Как, например, миграционное поведение молодых кукушек можно исчерпывающе объяснить на языке ДНК и синтеза белка? Очевидно, удовлетворительное объяснение потребует большего, нежели демонстрации того, что соответствующие гены, содержащие определённые последовательности оснований в ДНК, были необходимы для этого поведения или что поведение кукушат зависело от электрических импульсов в нервах; оно потребует некоторого понимания связей между специфическими последовательностями оснований в ДНК, нервной системой птиц и их миграционным поведением. На сегодняшний день, такая связь может быть определена лишь с помощью всё тех же неуловимых сущностей, которые «объясняют» все явления морфогенеза: витальных факторов, морфогенетических полей или генетических программ.
Более того, понимание поведения предполагает понимание морфогенеза. Так, даже если бы все особенности поведения относительно простого низшего животного, скажем, червя нематоды, могли быть детально объяснены на языке «электрических схем» (wiring) и физиологии е- го нервной системы, всё же было бы проблематично выяснить, каким образом нервная система с таким характерным расположением «электрических проводов» возникла в этом животном в процессе его развития.
1.4. Эволюция
Задолго до того, как была открыта менделевская генетика12, многие отдельные виды и породы домашних животных и растений были выведены путём селективного разведения (скрещивания). Нет причин сомневаться в том, что аналогичное развитие рас и видов происходит в дикой природе под влиянием скорее естественного, чем искусственного отбора. Неодарвинистская теория эволюции утверждает, что она может объяснить этот тип эволюции с помощью случайных мутаций. Но даже в рамках механистического мышления невозможно согласиться с тем, что этот тип маломасштабной или микроэволюции в пределах вида может объяснить происхождение самих видов, родов, семейств и высших таксономических единиц13. Одна научная школа полагает, что вся крупномасштабная или макроэволюция может быть объяснена с помощью длительных процессов микроэволюции14; другая школа это отрицает и утверждает, что большие скачки происходят внезапно в процессе эволюции15. Но, хотя в механистической биологии мнения об относительной значимости в макроэволюции множества малых или нескольких больших мутаций расходятся, существует общее согласие в том, что все эти мутации случайны и что эволюция может быть объяснена с помощью комбинации случайных мутаций и естественного отбора.
Однако эта теория всегда останется спекулятивной. Свидетельства об эволюции, получаемые в палеонтологии на основании исследования ископаемых, всегда будут давать пищу для разнообразных интерпретаций. Например, оппоненты механистической теории могут доказывать, что эволюционные изменения нельзя полностью объяснить случайными событиями, но что они обусловлены действием творческого принципа, непризнаваемого механистической наукой. Более того, влияния отбора, которые возникают в результате поведения и свойств самих живых организмов, можно рассматривать как зависящие от внутреннего организующего фактора, который по существу своему не механистичен.
Таким образом, проблема эволюции не может быть решена окончательно. Виталистические и организмические теории непременно включают экстраполяцию виталистических и организмических идей, точно так же как неодарвинистская теория содержит экстраполяцию механистических идей.
Это неизбежно; эволюция всегда будет рассматриваться в свете идей, возникших ранее на основе других предпосылок.
1.5. Происхождение жизни
Эта проблема так же неразрешима, как и проблема эволюции, и по тем же самым причинам. Во-первых, никогда нельзя точно знать, что происходило в отдалённые времена. Вероятно всегда будет изобилие спекуляций относительно обстоятельств появления жизни на Земле. Имеющиеся представления по этому вопросу следующие: земное происхождение жизни в Первичном Бульоне; заражение Земли микроорганизмами, умышленно принесёнными сюда на космическом корабле разумными существами с планеты, находящейся в другой солнечной системе16; эволюция жизни на кометах, содержащих органические вещества, возникшие из межзвёздной пыли17.
Во-вторых, даже если бы условия, при которых возникла жизнь, могли быть известны, это не пролило бы света на саму природу жизни. Если бы, предположим, можно было продемонстрировать, что первые живые организмы возникли из неживых агрегатов химических соединений или в результате «гиперциклов» химических процессов18, в Первичном Бульоне, это всё равно не доказывало бы, что такие процессы полностью механистичны. Органицисты всегда могли бы возразить, что возникли новые организмические свойства, а виталисты — что витальный фактор вошёл в первую живую систему именно тогда, когда она впервые появилась. Те же аргументы были бы пригодны, даже если бы живые организмы могли быть когда-либо синтезированы искусственным путём из химических веществ в пробирке.
1.6. Недостатки физического объяснения
Механистическая теория постулирует, что все явления жизни, включая поведение человека, могут быть в принципе объяснены на языке физики. Помимо любых споров, которые могут возникнуть из-за применения отдельных теорий современной физики или из-за конфликтов между ними, этот постулат проблематичен по крайней мере по двум фундаментальным причинам.
Во-первых, механистическая теория могла бы быть применима только, если бы физический мир был причинно замкнут. В отношении человеческого поведения это осуществилось бы в том случае, если бы ментальные состояния либо вовсе были нереальны, либо были бы в некотором смысле идентичны физическим состояниям тела, или протекали бы параллельно с ними, или представляли бы собой их эпифеномены19. Но, с другой стороны, если ум считать нефизическим и всё же причинно действенным, способным взаимодействовать с телом, тогда поведение человека не могло бы быть полностью объяснимо в физических терминах. Вероятность того, что ум и тело взаимодействуют, подтверждается доступными (экспериментальными) свидетельствами20: в настоящее время на основе эмпирических данных не может быть сделан определённый выбор между механистической теорией и теорией взаимодействия; с научной точки зрения вопрос остается открытым. Поэтому очевидно, что по крайней мере человеческое поведение даже в принципе не может быть полностью объяснено на языке физики.
Во-вторых, попытка объяснить умственную деятельность, используя физические термины, предполагает неизбежную «кругообразность», связанную с тем, что сама наука зависит от умственной деятельности21. Эта проблема стала очевидной в современной физике в связи с ролью наблюдателя в процессе физического измерения; принципы физики «даже не могут быть сформулированы без обращения (хотя, в некоторых вариантах, лишь подразумевающегося) к впечатлениям — и следовательно, к умам наблюдателей» (Б.Д’Эспаньа22). Таким образом, поскольку физика предполагает участие умов наблюдателей, эти умы и их свойства не могут быть объяснены на языке физики22.
1.7. Психология
В психологии, науке об уме, проблему отношения между умом и телом можно избежать, игнорируя существование ментальных состояний. Таков подход школы бихевиоризма, которая сосредотачивает своё внимание исключительно на объективно наблюдаемом поведении23. Но бихевиоризм не является проверяемой научной гипотезой — это методология. И его никак нельзя считать единственным подходом в психологии24.
Другие школы психологии приняли более прямой подход, используя в качестве первичных данных субъективный опыт. Для целей нашего обсуждения нет нужды рассматривать все существующие школы и системы; достаточно будет одного примера, чтобы показать трудности биологии, возникающие при применении психологической гипотезы, разработанной в попытке объяснить эмпирические наблюдения. Психоаналитические школы постулируют, что многие аспекты поведения и субъективного опыта зависят от подсознательного или бессознательного ума. Чтобы объяснить факты, имеющие место в момент пробуждения или во сне, бессознательный ум следует наделить качествами, совершенно отличными от качеств любой известной механической или физической системы. В концепции К.Г.Юнга бессознательный ум даже не ограничен умами идивидуумов, но представляет собой субстрат, общий для всех человеческих умов, или коллективное бессознательное: «В дополнение к нашему непосредственному (immediate) сознанию, которое имеет строго индивидуальную природу и которое, как мы полагаем, является единственной эмпирически изучаемой психикой (даже если мы присоединяем к нему в качестве приложения персональное бессознательное), существует вторая система психики, которая имеет коллективную, универсальную и безличностную природу и которая одинакова во всех индивидуумах. Это коллективное бессознательное не развивается индивидуально, но наследуется. Оно состоит из предсуществующих форм, архетипов, которые могут стать сознательными лишь вторично (опосредованно) и которые дают определённую форму данному психическому содержанию»26.
Юнг стремился объяснить наследование коллективного бессознательного физическими причинами, предполагая, что архетипные формы «присутствуют в семенной плазме»27. Но весьма сомнительно, что что-либо, обладающее свойствами архетипных форм, может быть наследуемо химически в структуре ДНК или в какой-либо иной физической или химической структуре клеток спермы или яйцеклетки. В действительности, идея коллективного бессознательного не очень осмыслена с точки зрения современной механистической биологии, каковы бы ни были её преимущества как психологической теории.
Однако нет априорных причин полагать, что психологические теории должны быть непременно ограничены рамками механистической позиции; они лучше вписываются в контекст теории взаимодействия (интеракционизма). Умственные феномены необязательно должны зависеть от физических законов; скорее, они должны следовать своим собственным законам.
Различие между механистическим и интеракционистским подходами можно проиллюстрировать, рассматривая проблему памяти. Согласно механистической теории, запоминаемое должно каким-то образом сохраняться в мозгу. Но с позиций интеракционизма, прошлые состояния ума могут влиять на настоящие его состояния непосредственно, способом, который не зависит от сохранённых следов физической памяти28. Если бы это было так, то поиски следов физической памяти в мозгу неизбежно были бы бесплодными. И хотя было выдвинуто несколько различных механистических теорий, рассматривающих, например, рефлектирующие циклы нервной активности; изменения синаптических связей между нервами; специфические молекулы РНК, — нет убедительных свидетельств, что любые из предложенных механизмов действительно могут объяснить, что такое память29.
Если память не сохраняется физически в мозгу, тогда некоторые виды памяти не обязательно должны быть ограничены умами индивидуумов; представление Юнга о наследуемом коллективном бессознательном, содержащем архетипные формы, можно интерпретировать как вид коллективной памяти.
Такие предположения, защищаемые в рамка- х интеракционизма, кажутся бессмысленными с механистической точки зрения. Но механистическая теория не может приниматься на веру; на сегодняшний день сама мысль, что все психологические феномены в принципе объяснимы в физических терминах, является не более чем спекуляцией.
1.8. Парапсихология
Во всех традиционных обществах рассказываются истории о мужчинах и женщинах, имеющих способности, кажущиеся сверхъестественными, такие способности засвидетельствованы и всеми религиями. В самых разных местах мира люди утверждают, что различные паранормальные способности целенаправленно развиваются такими системами, как шаманизм, колдовство, тантрическая йога и спиритуализм. И даже в современном западном обществе постоянно сообщается о фактах необъяснимых явлений — телепатии, ясновидении, предсказаниях, памяти о прошлых жизнях, домовых (призраках), полтергейстерах, психокинезе и так далее.
Очевидно, что это область, в которой распространены предрассудки, мошенничество и легковерие. Но возможность, что кажущиеся паранормальные события действительно происходят, не может быть исключена; ответ на вопрос может быть найден лишь после исследования явления. Научное исследование явлений, которые считаются паранормальными, продолжается уже почти столетие. Хотя в этой области исследований учёные обнаружили много случаев мошенничества и нашли, что некоторые явления, кажущиеся паранормальными, на самом деле могут быть объяснены нормальными причинами, остаётся множество свидетельств, которые, по-видимому, не могут быть объяснены на основе каких-либо из известных физических принципов30‘. Более того, многочисленные эксперименты, поставленные с целью проверки так называемого экстрасенсорного восприятия или психокинеза, дали положительные результаты с шансами против случайного совпадения, равными тысячам, миллионам или даже биллионам против одного31.
Поскольку эти явления не могут быть объяснены с помощью известных законов физики и химии, с механистической точки зрения, они и не должны происходить32. Но если они происходят, здесь возможны теоретические подходы двух типов. Первый основывается на исходном предположении, что такие явления зависят от законов физики, доселе неизвестных; второй — что они зависят от нефизических причинных факторов или от принципов, связывающих и те, и другие33. Большинство гипотез второго типа, выдвинутых к настоящему времени, были созданы в рамках интеракционизма. Несколько недавно предложенных гипотез основаны на формулах квантовой теории, включающих «невидимые переменные» или «разветвляющиеся вселенные», и постулируют, что ментальные состояния играют роль в определении результатов вероятностных процессов физического превращения34. Как неопределённость таких теоретических построений, так и неуловимость относящихся к ним феноменов являются причиной того, что исследования в области парапсихологии прогрессируют очень медленно. Это, в свою очередь, усиливает тенденцию многих механистически мыслящих биологов игнорировать или даже отрицать свидетельства, показывающие, что эти явления действительно существуют.
1.9. Выводы
Это краткое рассмотрение широко известных проблем биологии не оставляет больших надежд на то, что все они могут быть решены исключительно на основе механистического подхода. В случае морфогенеза и поведения животных вопрос следует считать открытым. Но проблемы эволюции и происхождения жизни не решаемы как таковые и не могут помочь сделать выбор между механистической и другими теориями жизни. Механистическая теория встречает серьёзные философские затруднения в связи с проблемой ограниченности физического объяснения; в отношении психологии она не имеет явных преимуществ перед теорией интеракционизма; и она противоречит несомненным фактам в случае парапсихологических феноменов.
В то же время, хотя интеракционизм может быть привлекательной альтернативой в области психологии и парапсихологии, он имеет серьёзный недостаток — создаёт пропасть между психологией и физикой. Более того, остаётся непонятным, каковы его биологические приложения. Если взаимодействие ума и тела влияет на поведение человека, то что можно сказать о поведении животных? А если нефизический причинный фактор участвует в управлении поведением животных, может ли он также играть роль в контролировании явления морфогенеза? В этом случае, следует ли его рассматривать как фактор того же рода, что и предлагаемый виталистическими теориями морфогенеза? Если да, то в каком смысле виталистический фактор, контролирующий эмбриональное развитие, напоминает ум человека?
Таким образом, теория интеракционизма, рассматриваемая в общебиологическом контексте, создаёт больше проблем, чем решает. И она, по-видимому, не позволяет сделать какие- либо проверяемые предсказания, кроме того, что она допускает возможность парапсихологических феноменов.
Организмический подход в его настоящем состоянии также страдает тем недостатком, что не предлагает новых путей эмпирического исследования; он не предлагает экспериментальной биологии почти ничего, кроме весьма неопределённой терминологии. При столь ненадёжных альтернативах биологические исследования должны продолжать следовать механистическому подходу, несмотря на его недостатки. На этом пути, по крайней мере, что-то можно выяснить, даже если главные проблемы биологии останутся нерешёнными. Но, хотя на короткий период это представляется единственно возможным способом действия, всё же, если заглянуть в будущее, кажется разумным задать вопрос, может ли альтернативный подход развиваться конкретно и последовательно и давать проверяемые предсказания? Если такая теория может быть сформулирована, проблема морфогенеза представляет, по-видимому, наиболее доступную отправную точку.
Перспективы создания более совершенных вариантов механистической, виталистической и организмической теорий морфогенеза обсуждаются в следующей главе.
Перевод с английского к.х.н. Е.М.Егоровой
(Продолжение следует)
Если ориентироваться на критерии роста, связанные с увеличением в процессе развития общей массы организма, то сразу же возникают затруднения при сопоставлениях не только представителей разных классов животных, но даже в пределах одного вида. Например, у млекопитающих рост особи прекращается при достижении определенного возраста и размеров, тогда как у рыб он продолжается в течение всей жизни. Среди представителей одного вида встречаются линии, расы и породы, которые существенно отличаются по темпам роста и массе тела. В частности, собаки крупных пород в течение равного периода онтогенеза в десятки раз превосходят по росту своих собратьев мелких пород. Точно так же отличаются по размерам тела и органов лошади-тяжеловозы и шотландский пони. Породные и видовые различия в размерах тела и органов, как правило, связаны с количеством составляющих их клеточных единиц (без заметных колебаний в размерах самих клеток) и являются довольно консервативными. Если, например, зачаток органа из эмбриона мыши пересадить в эмбрион слона, то рост пересаженного органа и количественное представительство составляющих его клеток будут лимитированы размерами органа мыши. Не без оснований поэтому ростовые процессы связывают с генетической конституцией особи, линии, вида (Бодемер, 1971; Goss, 1972а, b; Eiben, 1976; McKeown е. а.,1976). Однако известны случаи, когда при идентичной генетической детерминированности показатели роста в онтогенезе изменяются из-за нарушений в эндокринной системе. Поэтому в регулировании ростовых процессов на различных этапах онтогенеза важное значение имеет гормональный статус матери и развивающегося плода.